
ҶОјҡ°ыЮDдӣҪТКҫДcөАҫъИәјУЦШЦ¬·ҫёО


СРҫҝІДБП
јјРgВ·ҫҖ
ІҪуE1ЈәHFHCпӢКіХTҢ§өДNASHРЎКуёОЕKЦРҙжФЪҙЩСЧРФBјҡ°ыөДҫЫјҜәН»о»ҜЈ¬¶шBјҡ°ыИұПЭРЎКуөДёОЕKСЧ°YәНАwҫS»ҜөГөҪёДЙЖ
ІҪуE2ЈәMyD88КЗBјҡ°ыФЪNASHЯ^іМЦРҙЩЯMёОЕKСЧ°YЎўАwҫS»ҜәН“pӮыЛщұШРиөДЈ»
ІҪуE3ЈәNASHЦРөДBјҡ°ы»о»ҜКЬөҪBјҡ°ыКЬуwРЕМ–Ј¬МбКҫBјҡ°ы»о»ҜХыәПБЛМмИ»ГвТЯәНЯm‘ӘРФГвТЯҷCЦЖ;
ІҪуE4ЈәТЖЦІNASH»јХЯјSҫъФцјУБЛКЬуwРЎКуёОЕKЦРөДBјҡ°ы·eАЫәН»о»ҜЈ¬ҸД¶шјУ„ЎNASHЈ¬МбКҫДcөАҫъИәТтЛШтҢ„УБЛBјҡ°ыФЪNASHЦРөДЦВІЎРФЎЈ
СРҫҝҪY№ы
һйБЛҙ_¶ЁёОғИBјҡ°ыФЪNASHЯMХ№ЦРөДЧчУГЈ¬ЧчХЯУГHFHCпӢКіХTҢ§РЎКуNASHЈ¬Іў·ЦлxіцГвТЯјҡ°ыәуЧR„eЦчТӘөДГвТЯјҡ°ыИәЈ¬HFHCҪMөДBјҡ°ып@ЦшФцјУЎЈһйБЛФu№АBјҡ°ыЦРјҡ°ыТтЧУөДұнЯ_ЗйӣrЈ¬ЧчХЯ°l¬F·Ц„eФЪұы¶юҙјјЧГСҙЧЛбхҘЈЁPMAЈ©әНЦ¬¶аМЗЈЁLPSЈ©ҙМјӨәуЈ¬NASHРЎКуұнЯ_TNF-αәНIL-6өДBјҡ°ы”өБҝФцјУЎЈН¬•rЈ¬ҒнЧФNASHРЎКуөДBјҡ°ыТІп@Кҫіцјҡ°ыұнГжЦчТӘҪMҝ—ПаИЭРФҸНәПОп (MHC) IоҗәНIIоҗТФј°CD86өДЙПХ{Ј¬Я@ұнГч»о»ҜәНҝ№ФӯіКЯfДЬБҰФцјУЎЈТФЙПҪY№ыұнГчЈ¬NASHРЎКуЦРёОғИBјҡ°ы·eАЫЈ¬ІўбҢ·ЕҙЩСЧјҡ°ыТтЧУТФј°іКЯfҝ№ФӯөДДЬБҰФцјУЎЈ
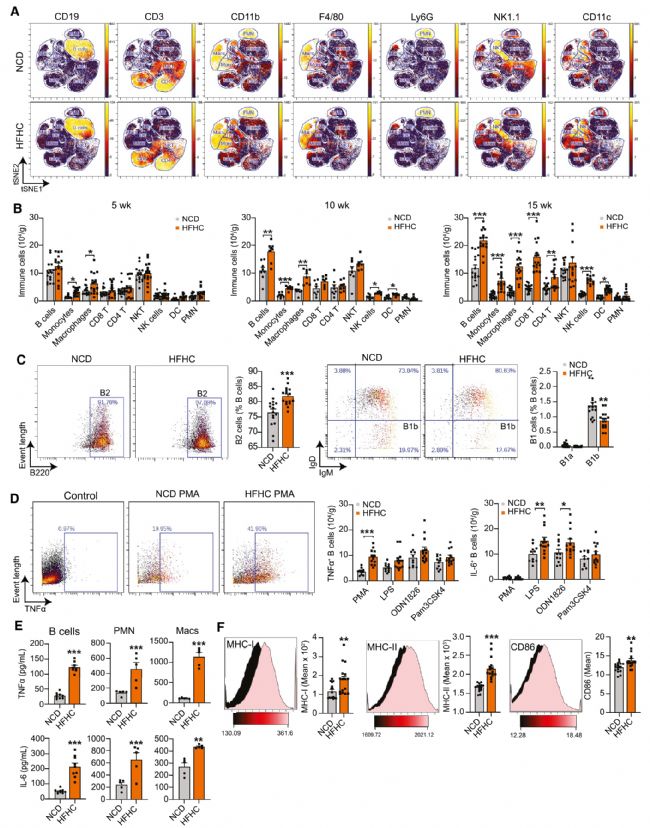
ҲD1 ҙЩСЧРФBјҡ°ыФЪNASHРЎКуөДёОЕKЦР·eАЫ
2. NASHРЎКуёОғИBјҡ°ып@КҫҙЩСЧ»щТтЧV
һйБЛёьәГөШБЛҪвNASHИзәОёДЧғBјҡ°ыөДЮDдӣҪMЧVЈ¬ЧчХЯҢҰҒнЧФNCDәНHFHCёОЕKөДГвТЯјҡ°ыЯMРРБЛҶОјҡ°ыЮDдӣҪMңyРтЈ¬°l¬F24ӮҖCD45+ГвТЯјҡ°ыҙШЎЈЧчХЯЦчТӘкPЧўБЛBјҡ°ыҙШөД»щТтұнЯ_ЧVЈ¬°l¬FСЧ°Y»щТтIl1bЎўS100a8өИФЪHFHCёОЕKBјҡ°ыЦРұнЯ_ФцјУЎЈһйБЛтһЧCҶОјҡ°ыЮDдӣҪMҢҰBјҡ°ыұнЯ_ЧVөДҷzңyДЬБҰЈ¬ЧчХЯЯҖҢҰHFHCәНNCDРЎКуёОЕKBјҡ°ыЯMРРБЛіЈТҺRNA-seqЈ¬Н¬ҳУтһЧCБЛҺЧӮҖҙЩСЧ»щТтФЪHFHCёОЕKBјҡ°ыЦРЙПХ{ЎЈһйБЛҪвбҢЯ@Р©”ө“юөДЙъОпҢWТвБxЈ¬ЧчХЯК№УГ Ingenuity Pathway Analysis (IPA) Ғнҷzңy»щТтұнЯ_ДЈКҪЈ¬°l¬F…ўЕcСЧ°YәНTLRРЕМ–ӮчҢ§өДЙПУОХ{№қТтЧУөДјӨ»оЈ¬ТФј°IL10RAәНSOCS1өДТЦЦЖЈ¬ІўЗТё»јҜөҪ»оЬSөДСЧ°YНҫҸҪЎЈТФЙПҪY№ып@КҫNASHРЎКуөДBјҡ°ы»о»ҜәНСЧ°YұнРНФцјУЎЈ
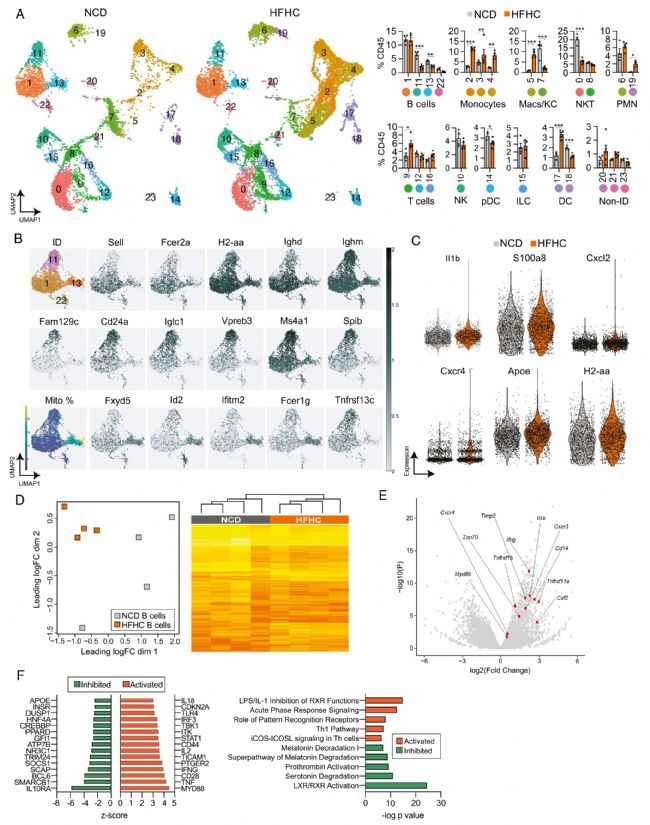
ҲD2 ёОғИBјҡ°ыФЪNASHЖЪйgп@КҫіцҙЩСЧ»щТтЧV
3. Bјҡ°ыИұПЭҝЙёДЙЖNASH
һйБЛҙ_¶ЁBјҡ°ыКЗ·сФЪNASHөД°lІЎҷCЦЖЦРЖрЦұҪУЧчУГЈ¬ЧчХЯФu№АБЛHFHCО№рBөДBјҡ°ыИұПЭμMTРЎКуөДNASHЯMХ№ЎЈHFHC μMTРЎКуп@КҫіцNASФu·ЦЭ^өНЈ¬ЗТЮDдӣҪM”ө“юұнГчЈ¬ФЪHFHC μMTРЎКуЦРЈ¬ҙЩСЧ»щТтөДұнЯ_п@ЦшҪөөНЈ¬Bјҡ°ы”өДҝп@ЦшңpЙЩЈ¬ІўЗТHFHC μMTРЎКуАwҫS»ҜГчп@ңpЙЩЈ¬АwҫS»ҜРОіЙ»щТтұнЯ_ҪөөНЈ¬¶шЯ@Р©Чғ»ҜФЪNCD WTәНNCD μMTРЎКуЦРІўОҙҷzңyөҪЈ¬Ф“ҪY№ыХfГчФЪHFHCО№рBәуЈ¬Bјҡ°ыҝЙТФТэЖрҙъЦxәНСЧ°YОЙҒyЎЈ
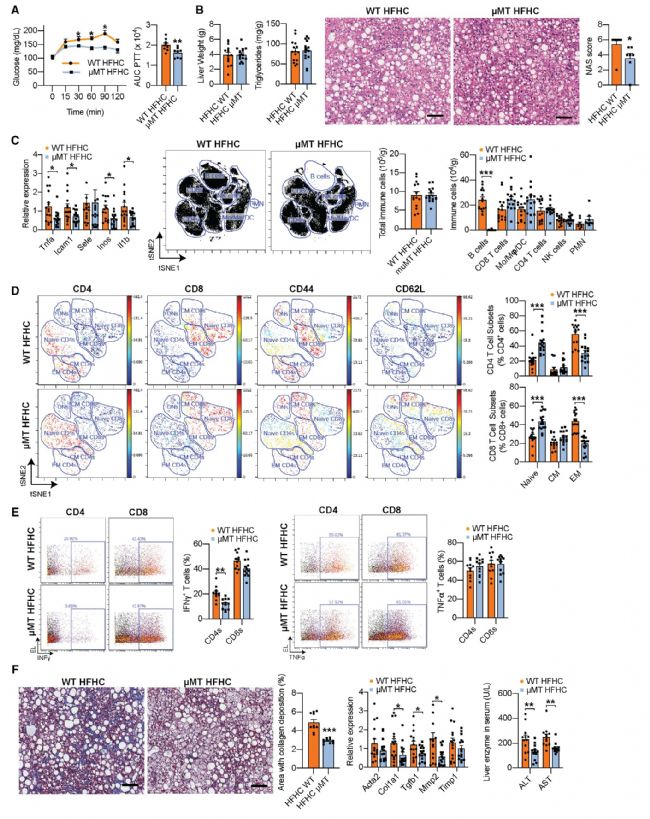
ҲD3 B јҡ°ыИұ·ҰҝЙёДЙЖNASHЖЪйgөДСЧ°YәНАwҫS»Ҝ
4. ҙЩСЧ»щТтMyD88ҪйҢ§NASHЦРBјҡ°ыөДҙЩСЧ№ҰДЬ
RNA-seq·ЦОцҙ_¶ЁMyD88КЗHFHCРЎКуёОBјҡ°ыЦРҙЩСЧ»щТтөДЙПУОХ{№қТтЧУЈ¬һйБЛСРҫҝBјҡ°ы№МУРөДMyD88РЕМ–ӮчҢ§КЗ·сУ°н‘NASHЯMХ№Ј¬ЧчХЯҳӢҪЁБЛBјҡ°ыМШ®җРФИұК§ MyD88 (B-MYD) өДРЎКуЈ¬HFHCО№рBәу°l¬FB-MYD HFHCРЎКуNASФu·ЦЭ^өНЎЈлmИ»B-MYD HFHCәНWT HFHCРЎКуЦ®йgөДҝӮ”өәНҒҶИәГвТЯјҡ°ыИәӣ]УРІо®җЈ¬ө«B-MYD HFHCРЎКуөДёОғИBјҡ°ыMHC-IәНMHC-IIңpЙЩЈ¬Я@ұнГчTLRҪйҢ§өДјӨ»оҪөөНБЛҝ№ФӯөДіКЯfДЬБҰЎЈТФЙПҪY№ыұнГчЈ¬Bјҡ°ыРиТӘMyD88ҒнҙЩЯMNASHөДёОЕKСЧ°YЎўАwҫS»ҜәН“pӮыЎЈ
ҲD4 MyD88ҪйҢ§NASHЦРBјҡ°ыөДСЧ°Y№ҰДЬ
5. BCRҙМјӨјӨ»оёОғИBјҡ°ы
BCRЕcҝ№ФӯҪYәП®aЙъөДјӨ»оРЕМ–КЗBјҡ°ы»о»ҜөДөЪТ»РЕМ–Ј¬ҪУПВҒнЧчХЯМҪҫҝБЛФЪNASHЯ^іМЦРёОғИBјҡ°ыөДјӨ»оКЗ·сЕcBCRРЕМ–УРкPЈ¬ТтBCRРЕМ–ӮчҢ§ФЪNur77»щТтөДҝШЦЖПВХTҢ§ҫGЙ«ҹЙ№вө°°Ч (GFP) ұнЯ_Ј¬ЧчХЯҪoNur77-GFPРЎКуО№КіNCD»тHFHCЈ¬ҪY№ы°l¬FHFHC Nur77-GFPРЎКуөДGFP+ёОғИBјҡ°ыөДоlВКәН”өБҝп@ЦшФцјУЈ¬ұнГчФЪNASHЖЪйgөДјӨ»оЙжј°ёОЕKЦРөДBCRРЕМ–ЎЈФ“ҪY№ыұнГчЈ¬NASHЦРЈ¬Bјҡ°ыөД»о»ҜКЬөҪBCRРЕМ–ҙМјӨЎЈ
ҲD5 Bјҡ°ыНЁЯ^BCRұ»јӨ»о
6. ДcФҙРФОўЙъОпТтЧУҙЩЯMёОғИBјҡ°ы»о»Ҝ
һйБЛСРҫҝNAFLDПакPҫъИәК§Х{®aЙъөДјҡҫъ®aОпКЗ·сҙЩЯMёОғИBјҡ°ы»о»ҜЈ¬ЧчХЯҳӢҪЁБЛFMTРЎКуДЈРНЈ¬ЕcҪЎҝөFMTРЎКуПаұИЈ¬NAFLD FMTРЎКуұн¬Fіц”M—UҫъйTәНЧғРОҫъйTШS¶ИФцјУЈ¬Ф“ҪY№ыЕcNAFLD№©уwөДҫъИәұИАэҫЯУРёЯ¶ИПаЛЖРФЎЈЗТNAFLD FMTРЎКуNASФu·ЦВФУРФцјУЈ¬ёОғИBјҡ°ы”өБҝФцјУЈ¬ҝ№ФӯіКЯfәН№ІҙМјӨ·ЦЧУөДұнЯ_ФцјУЈ¬АwҫS»ҜЭpОўФцјУЎЈЯ@Р©”ө“юұнГчЈ¬ДcөАҫъИәЦРЕcNAFLDПакPөДЧғ»ҜЧгТФХTҢ§ёОЕKЦРBјҡ°ыөД·eАЫәН»о»ҜЈ¬Н¬•rҢўёОЕKІЎАнҢWөДёчӮҖ·ҪГжҸДИЛоҗ№©уwӮчЯfҪoКЬуwРЎКуЎЈ
ҲD6 ДcФҙРФОўЙъОпТтЧУҙЩЯMёОғИBјҡ°ы»о»Ҝ
ҪYХ“

- Л®Щ|·ЦОцПөҪyҢҚтһКТФOУӢТҺ„қФ”Ҫв
- ҪТКҫРЎКуЕЯМҘДёФҙ-әПЧУЮD“QЯ^іМЦРөДRNA m6A„У‘B
- ҶОјҡ°ыј°ҝХйg¶аҪMҢWҪТКҫAPOE4ФЪЛҘАПЎўЙсҪӣСЧ°YәН°ў –ЖқәЈД¬°YөДҷCЦЖ
- АыУГACSL4ҙЩЯMЙсҪӣСЧ°YРВҷCЦЖөДСРҫҝ
- Int J Mol SciЈә¶аҪMҢWҪТКҫMCT2ПЮЦЖ·О°©РЎКуДЈРНЦРөДД[БцЙъйL
- ДcөАҫъИә-°РЖч№Щ¶аҪMҢWЈә16sәН·З°РҙъЦxҪMҢWҷzңy‘ӘУГУЪЛ®МKМЗСРҫҝ
- ЦРбtуwЩ|ҢWөДЙъАнҢW»щөAҪйҪB
- ёЙШӣ·ЦПн--ёЯНЁБҝңyРтФӯКј”ө“юЙПӮчҪМіМ
- БҝңКSPR»ҘЧчҷzңy·ю„ХЈәРФғrұИһйәОИзҙЛЦ®ёЯЈҝ
- әјЦЭҠWКўҳsвЯЎ°2023ЦРҮшДк¶ИғһЯx№НЦчЎұ
- ЗЪПиҝЖҢWғxЖчКЪҷа·ю„Х№ӨіМҺҹЕаУ–•юЧhФЪьSЙҪнҳАыЕeЮk
- ЦРҝЖРВЙъГь3ФВ°lұнОДХВ37ЖӘЈ¬ЖҪҫщУ°н‘ТтЧУ8.12·ЦЈЎ
- ЦРҝЖРВЙъГьHL2400ёЯНЁБҝ°РПтЦ¬Щ|ҪMЙэјү-"Ц¬"өГ“нУР
- ҙәЕҜ»Ёй_,ёР¶чПа°й--ЦРҝЖРВЙъГьЙо¶ИҝХйgҙъЦxҪMҙуҙЩ
- ЦРҝЖРВЙъГь2023APTоIУтҪвӣQ·Ҫ°ёИ«ҮшСІЦvИ«ГжҶў„У
- ЦРҝЖРВЙъГь"СӘТәҙъЦxҪM-CellөИёЯ·ЦОДХВТ»Чч·ЦПн•ю"


