
Îŧ°ûŪDäŊMyĐōŊŌĘžN7-ŧ×ģųøBÜÕËáŖ¨m7GŖŠtRNAĐŪīpČõĩÄÖ²ĄCĀí

ÆÚŋ¯ŖēProceedings of the National Academy of Sciences
Ķ°íŌō×ĶŖē9.4
˛ŽēĀŧŧĐgˇūÕŖēÎŧ°ûēËŪDäŊMyĐōĄĸ˛Ž®ŧ°ûēˡÖëxÔŠēĐ
ŅĐžŋąŗž°
ąŗž°Ŗēm7Gŧ×ģųŪDŌÆøÍēĪķwŊMˇÖWDR4ĩÄÍģ×ÔÚÉņŊĐÔŧ˛˛ĄģŧÕßÖĐŗŖŌŖŦĩĢCÖÆ˛ģÃ÷ĄŖ
ŋÆWî}ŖēWDR4Õ{ŋØÉņŊŧ˛˛ĄĩġÖ×ĶCÖÆŖģÔCÖÆÔÚÖίÖĐĩÄÔÚĒĶÃrÖĩĄŖ
ō˛ÄÁĪŖēC57BL/6NĐĄĘķĄĸČËîiPSCÕT§ˇÖģ¯ŗÉĩÄÉņŊÔĒŧ°ûĄŖ
ąíĐÍŖēÔ´×Ô˛ĄČËĩÄWDR4ÍģקÖÂĐĄĘķĖØļ¨îĐÍÉņŊÔĒëSrégÍËģ¯ŖŦßMļø§ÖÂĐĄĘķÉņŊĪĩŊy°lĶũÎÉyĄŖ
CÖÆŖē
1. WDR4ücÍģ×ŌÖÖÆWDR4ēÍMETTL1ÖŽégĩÄģĨ×÷ŖŦĘšĩÃMETTL1ĩ°°×ŧĶËŲŊĩŊâŖģ
2. Wdr4Íģ×ĘštRNA m7GĐŪīĪÂÕ{ŖŦßMļøĪÂÕ{mTORͨ¡ĪāęPmRNAĩġ×gŖŦŌũ°lČŲ|žWĒŧ¤ŖŦÉĪÕ{ÉņŊÔĒŧ°ûĩōÍöŖģ
ĒĶÃŖē
1. TUDCAĖĀíŋÉŌÔī@ÖøžŊâWdr4ÍģקíĩÄÉņŊÔĒËĀÍöŖŦĖáÉũWdr4Íģ×ĐĄĘķĩÄÉņŊÔĒšĻÄÜŖģ
2. AAV˛ĄļžŪD§ģØŅaŌ°ÉúĐÍWDR4ŖŦÄÜōģÖÍĐĄĘķ´ķÄXMETTL1ĩ°°×ŧ°tRNA m7GĐŪīËŽÆŊŖŦĻWdr4Íģ×ĐĄĘķžßĶĐéLЧÖÎ¯Đ§šûŖģ
3. ĻžßĶĐÅR´˛˛ĄČËWDR4Íģ×ĩÄČËîiPSCÕT§ŗÉĩÄÉņŊÔĒŧ°ûßMĐĐTUDCAĖĀíēÍWDR4ģØŅaŖŦŧ°û´æģîÂĘēÍšĻÄÜĐÔžųĶĐī@ÖøĖáÉũĄŖ
˛ŽēĀŽaÆˇ
Îŧ°ûēËŪDäŊMyĐōĄĸŪDäŊMyĐōĄĸTRAC-seqĄĸļāēËĖĮķwyĐō
Ŗ¨Îŧ°ûēÍŪDäŊMyĐōˇūÕĶɲŽēĀÉúÎīĖᚊŖŠ
ŅĐžŋŊYšû
1. WDR4ÍģקÖ°lĶũĄĸß\ĶēÍÖĮÁĻČąĪŨ
ÅR´˛ÉĪŖŦģŧÕßŗŖŌĩÄWDR4Íģ×ĘĮ170ÎģĩÄžĢ°ąËáŖ¨RŖŠÍģ×ŗÉÁÁ°ąËáŖ¨LŖŠŖŦ×÷Õßͨß^Wdr4ģųŌōĩÄļ¨ücÍģ×ŖŦʚХĘķWDR4 215RÍģ×éLŖŦĶÉ´ËŊ¨ŗöWDR4ÅR´˛Íģ×ĩÄĐĄĘķÄŖĐÍŖ¨Wdr4R21L/R215LŖŦĪ¡Q“Íģ×ĐĄĘķ”ŖģFig. 1A-CŖŠĄŖĪāąČĶÚŌ°ÉúĐÍĐĄĘķŖ¨Wdr4+/+ŖŠŖŦÍģ×ĐĄĘķŗöŦFĀÖØĩÄ°lĶũßtžŖŦķwÖØĄĸķwĐÍēÍ´æģîrégī@ÖøŊĩĩÍŖ¨Fig. 1D-GŖŠĄŖÁíŌģˇŊÃæŖŦ×÷ÕßŌ˛ĻŌ°ÉúĐÍēÍÍģ×ĐĄĘķĩÄß\ĶÆŊēâÄÜÁĻēÍÕJÖĒÄÜÁĻßMĐĐÁËzyēÍąČŨ^ŖŦ°lŦF8ÖÜũgĩÄÍģ×ĐĄĘķÔÚÉĪĘöˇŊÃæžų´æÔÚī@ÖøĩÄČąĪŨŖŦļø4ÖÜũgĐĄĘķt]ĶĐī@Öø˛îŽŖ¨Fig. 1H-PŖŠĄŖ
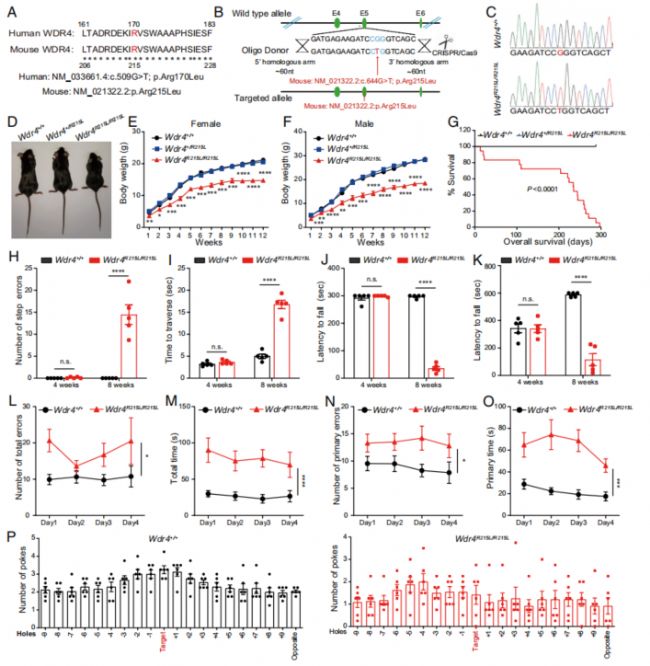
Fig. 1. Wdr4Íģ×ŌũÆđ°lĶũĄĸĶ×÷ēÍÖĮÁĻČąĪŨ
2. WDR4ücÍģקÖÂÉņŊÔĒŧ°ûËĀÍöēÍĐĄî^ģûĐÎ
ČįÍŦÅR´˛ÉĪĩÄWDR4ÍģײĄČËūŗöŦFĐĄî^ģûĐÎŖŦ8ÖÜũgĩÄÍģ×ĐĄĘķĪāŨ^ĶÚŌ°ÉúĻÕÕŊMŖŦÄX˛ŋŗß´įēÍÖØÁŋÉĪŌ˛ī@ÖøŊĩĩÍŖŦ´ķÄXƤĶoī@Öø˛îŽŖŦĩĢĐĄÄXÎŽŋsĀÖØŖŦ°éĶĐĐĄÄXÖĐĩōÍöŧ°ûĩÄŋĩÄī@ÖøÉĪÕ{Ŗ¨Fig. 2A-H, L-NŖŠĄŖŋŧ]ĩŊÍģ×ĐĄĘķÔÚ4ÖÜũg]ĶĐĐÎBĄĸÕJÖĒēÍĐĐéÉĪĩÄī@ÖøČąĪŨŖŦËųŌÔ×÷ÕßŌ˛Ļ2ÖÜÖÁ8ÖÜÖŽégĩÄļāÄęũgrégücĩÄĐĄĘķĐĄÄXßMĐĐÁËļ¨ÁŋˇÖÎöŖŦŋÉŌÍģ×ĐÍĐĄĘķĩÄĐĄÄXÎŽŋsąíĐÍÔÚ5ÖÜŖ¨Day 35ŖŠēķé_Ęŧī@ŦFŖŦÖ÷ŌĒķwŦFÔÚĐĄÄXƤĶÖĐIGLŖ¨internal granule layerŖŦȲŋîwÁŖĶŖŠēÍMLŖ¨molecular layerŖŦˇÖ×ĶĶŖŠĩÄÃæˇeī@ÖøŊĩĩÍŖ¨Fig. 2I-KŖŠĄŖ
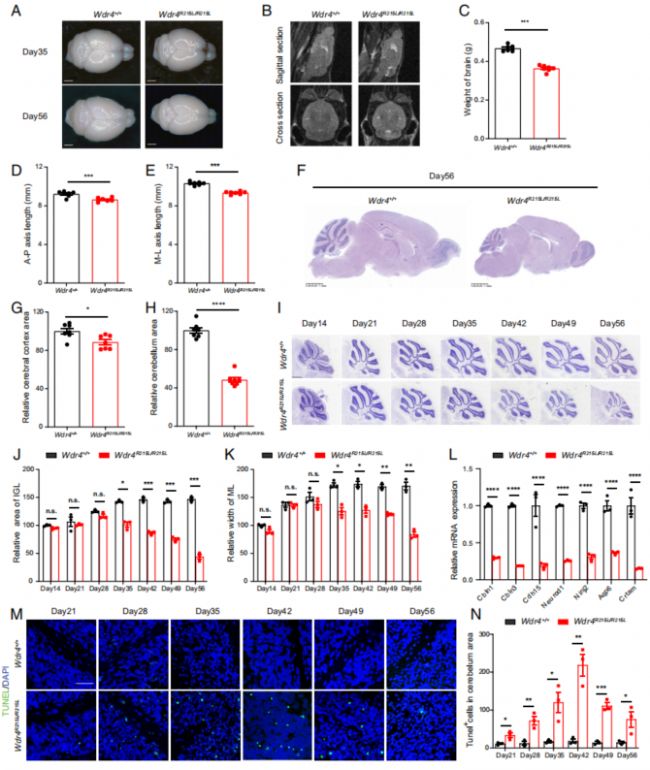
Fig. 2. WDR4ücÍģ×ŌũÆđÉņŊŧ°ûËĀÍöēÍĐĄî^ģûĐÎ
3. Wdr4Íģ×ĐĄĘķÄX˛ŋĖØļ¨îĐÍŧ°ûëSrégÍËģ¯
ÉĪĘöĐĄÄXÎŽŋsĩIJĄĀíß^ŗĖŌ˛°ĩĘžÁËMETTL1/WDR4ͨß^tRNA m7GÕ{ŋØÄX˛ŋ°lĶũÅcˇBĩÄCÖÆŌ˛ŋÉÄÜžßĶĐrĐōÉĪĩÄĖØÕ÷ĄŖŌō´ËŖŦ×÷ÕßÔÚŌ°ÉúĐÍĐĄĘķÖĐzyÁË1ČÕũgÖÁ8ÖÜũgÖŽégļārégücĩÄMETTL1ēÍWDR4ĩ°°×ËŽÆŊŖŦ°lŦFļūÕßÔÚ4ÖÜŖ¨Day 28ŖŠĮ°ëSrégÖđuÉũ¸ßŖŦÔÚDay 28×ķĶŌß_ĩŊˇåÖĩŖŦëSēķé_ĘŧÖđuĪÂŊĩŖ¨Fig. 3AŖŠĄŖ
éÁËÔÚÎŧ°ûËŽÆŊÉĪĖŊžŋWdr4ÍģקíĩÄĶ°íŖŦ×÷ÕßßxČĄ8ÖÜũgŖ¨Day 56ŖŠĐĄĘķßMĐĐÎŧ°ûēËyĐōŖ¨snRNA-seqŖģyĐōˇūÕĶɲŽēĀÉúÎīĖᚊŖŠŖŦͨß^ŌŅÖĒĩÄÄX˛ŋŧ°ûmarkerĻžÛîēķĩÄĩūßMĐĐÁË×ĸáŖŦ˛ĸĻÔÚĩÁŋÉĪÕŧŊ^´ķļāĩĩÄÉņŊÔĒŧ°ûßMĐĐÁËßMŌģ˛ŊĩÄĐÍŧˇÖÅc×ĸáŖ¨Fig. S2A, F, G; Fig. 3BŖŠĄŖÔÚÉņŊÔĒŧˇÖŗÉĩÄ14ČēÖĐŖŦSubcluster 0ÔÚÍģ×ĐĄĘķÖĐ×ēõĪûʧŖŦÔŧ°ûĐÍÖĐŖŦîwÁŖĶŧ°ûmarker Gabra6ēÍEtv1ĖØŽĐԸ߹íŖŦ°ĩĘžÁËWdr4ücÍģ×ĻĐĄÄXîwÁŖĶŧ°ûÍËģ¯ĩÄĖØŽĐÔÕ{ŋØŖ¨Fig. 3B-DŖŠĄŖGO¸ģŧ¯ˇÖÎöĩÄŊYšûī@ĘžSubcluster0ī@ÖøĪÂÕ{ĩIJîŽģųŌōŖ¨DEGsŖŠÖ÷ŌĒ¸ģŧ¯ĶÚÉņŊÔĒĐÎBŽŗŖēÍĐĄÄXĐÎBŽŗŖĩČͨ¡Ŗ¨Fig. 3E, FŖŠĄŖ
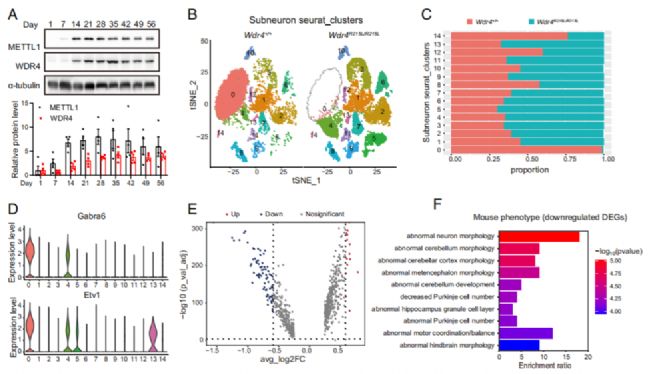
Fig. 3. Wdr4ücÍģ×ĐĄĘķÄX˛ŋŗöŦFĖØļ¨ŧ°ûîĐÍëSrégÍËģ¯
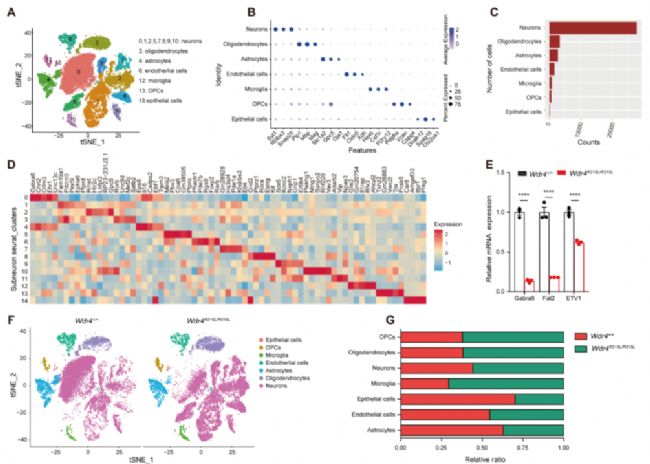
Fig. S2. 2ÔÂũgĐĄĘķÄX˛ŋsnRNA-seqˇÖÎö
4. Wdr4ücÍģ×ŌÖÖÆÆä×ÔÉíÅcMETTL1ĩÄģĨ×÷˛ĸŊĩĩÍMETTL1ˇļ¨ĐÔ
ŌÔÍųĩÄŅĐžŋķĩĀÁËMETTL1/WDR4´ßģ¯tRNAĩÄm7GĐŪīß@ŌģCÖÆŖŦĶÚĘĮ×÷ÕßŌÔ´ËéģųĩAŖŦßMŌģ˛ŊĖŊžŋMETTL1ēÍWDR4ÔÚÍģ×ĐĄĘķÖĐĩÄąíß_ĮérĄŖÄX˛ŋĩÄMETTL1ÔÚĐĄÄXÖи߹íļøÔÚÆäËû
^ĶōĩÄąíß_ÁŋŨ^ĩÍŖ¨ÅcĐĄÄXÎŽŋsąíĐÍĪāˇûŖŠŖŦWdr4ĩÄücÍģ×ī@ÖøĪÂÕ{ÁËtRNAĩÄm7GËŽÆŊēÍMETTL1ĩÄĩ°°×ËŽÆŊŖŦĩĢĘĮĻēķÕßĩÄŪDäËŽÆŊ]ĶĐī@ÖøĐÔĩÄĶ°íŖŦļøĩ°°×øķwŌÖÖÆŠMG132ĩÄĖĀíĘšĩÃWdr4Íģ×ĩÄÔ´úÉņŊŧ°ûÖĐMETTL1ĩÄĩ°°×ËŽÆŊī@ÖøÉĪÕ{ŖŦ°ĩĘžÁËWdr4ücÍģקÖÂÁËMETTL1ĩ°°×ĩÄŊĩŊâŖ¨Fig. 4A-EŖŠĄŖ
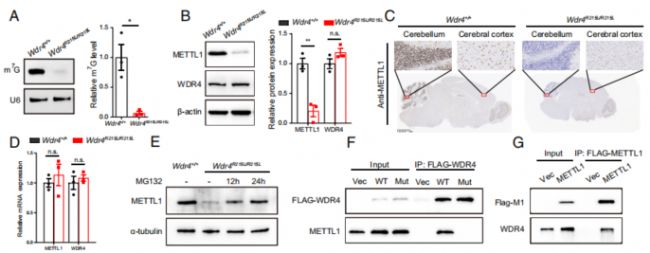
Fig. 4. Wdr4ücÍģ×ͨß^ĩ°°×ŊĩŊâŊĩĩÍMETTL1ˇļ¨ĐÔ
5. Wdr4ücÍģקÖÂm7G tRNAĐŪīĄĸmRNAˇ×gĪÂÕ{ŧ°ČŲ|žWĒŧ¤ÉĪÕ{
×÷ÕßĀûĶÃTRAC-seqßMŌģ˛ŊĖŊžŋWdr4ücÍģ×ĻÉņŊÍËģ¯ĩÄĶ°íŖŦēYßxĩŊÁËēŦĶĐm7GĐŪīĩÄtRNAģųĐō“RRRGGT”ŖŦ˛ĸ°lŦFÁËÍģ×ĐĄĘķÄX˛ŋtRNA m7GĐŪīËŽÆŊĩÄī@ÖøĪÂÕ{ŖŦēŦm7GĩÄtRNAËŽÆŊĪÂÕ{ĩĢ˛ģēŦm7GĩÄtRNAËŽÆŊ]ĶĐī@ÖøĐÔ×ģ¯Ŗ¨Fig. 5A-FŖŠĄŖāŅßĘÚËØōŊYšûī@ĘžŖŦÍģ×ĐĄĘķĩÄāŅßĘÚËØzČëī@ÖøŊĩĩÍŖŦŧ´ˇ×gËŽÆŊī@ÖøĪÂÕ{Ŗ¨Fig. 5GŖŠĄŖ×÷ÕßŊĶÖøĻŌ°ÉúĐÍēÍÍģ×ĐĄĘķĩÄÄX˛ŋßMĐĐÁËļāžÛēËĖĮķwyĐōŖ¨Polysome-seqŖŠŖŦŌÔĖŊžŋĐĄĘķÄX˛ŋmRNAĩġ×gЧÂĘŖŦŊYšûī@ĘžŖŦÔÚÍģ×ĐĄĘķÄX˛ŋˇ×gЧÂĘĪÂÕ{ĩÄģųŌō§ĶиüļāĩÄm7G tRNAy§ĩÄÃÜ´a×ĶŖŦĮŌTEĩÄ×ģ¯ÂĘÅcm7G ÃÜ´a×ĶŗöŦFĩÄîlÂĘĄĸĩÄŋŌÔŧ°ž´aĐōÁĐĩÄéLļČī@ÖøĪāęPŖ¨Fig. 5H, I; Fig. S4A-CŖŠĄŖ
TRAC-seqŊYšûÖĐŖŦˇ×gЧÂĘĪÂÕ{ĩÄģųŌōÖ÷ŌĒ¸ģŧ¯ÔÚŧ°ûĒŧ¤ĄĸÄX˛ŋ°lĶũēÍmTORĐÅĖͨ¡ÉĪŖŦļøRNA-seqĩÄŊYšûÖĐĶÖ¸ģŧ¯ĩŊÁËÉĪÕ{ĩÄŧ°ûĒŧ¤Í¨ÂˇŖŦŧ´ŧ°ûĒŧ¤ĪāęPģųŌōÔÚŪDäËŽÆŊÉĪĩÄÉĪÕ{ē͡×gËŽÆŊÉĪĩÄĪÂÕ{Ŗ¨Fig. 5J-OŖŠĄŖ´ËÍâWdr4ÍģקÖÂÁËČŲ|žWĒŧ¤ĪāęPģųŌōŪDäËŽÆŊÉĪĩÄÉĪÕ{ŧ°Ôͨ¡ÖĐęPæIˇÖ×ĶIRE1ēÍPERKĩÄÁ×Ëáģ¯ËŽÆŊÉĪÕ{ŖŦļøŊĩĩÍPERKÁ×Ëáģ¯ËŽÆŊtūÍŦrĘšĩôŲĩōÍöĩ°°×BaxēÍBimĩÄËŽÆŊī@ÖøĪÂÕ{Ŗ¨Fig. 5P, Q;Fig. S5ŖŠĄŖÁíÍâŖŦĶÉĶÚÎÄĢIķĩĀß^ČŲ|žWĒŧ¤ÅcĐĄÄzŲ|ŧ°ûŧ¤ģîÖŽégĩÄęPÂŖŦËųŌÔ×÷ÕßŌ˛Ļ´ËßMĐĐÁËō×CŖŦ°lŦFIbalęĐÔĩÄĐĄÄzŲ|ŧ°ûÔÚDay 35ÖŽēķĩÄÍģ×ĐĄĘķĐĄÄXÖĐŋėËŲUŖŦÅcÍģ×ĐĄĘķ×ÔDay 35é_ĘŧŗöŦFĐĄÄXÎŽŋsĩÄąíĐÍŌģÖÂŖ¨Fig. 5RŖŠĄŖ
žCÉĪŖŦ×÷Õß×CÃ÷ÁËĐĄĘķÄX˛ŋĩÄWdr4ücÍģ×ūĪÂÕ{tRNAÔÚĪāĒÎģücĩÄm7GĐŪīēÍmRNAˇ×gŖŦÉĪÕ{ŧ°ûĒŧ¤ēÍĩōÍöĄŖ
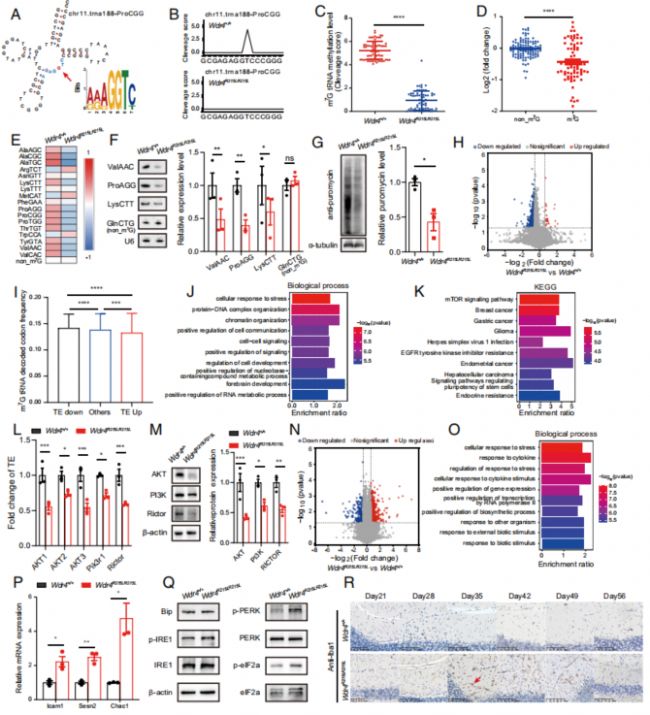
Fig. 5. Wdr4ücÍģ×ŌũÆđtRNA m7GĐŪīĪÂÕ{ŖŦmRNAˇ×gËŽÆŊĪÂÕ{ēÍČŲ|žWĒŧ¤ÉĪÕ{
6. ģØŅaōŧ°ÅR´˛ĒĶÃĮ°ž°
ęPĶÚģØŅaōĩIJŋˇÖŖŦ×÷ÕßŅĐōußMĩØÔOĶÁËČũˇN˛ģÍŦĩÄģØŅaōŖē
Ę×ĪČĘĮĀûĶÃFDAŌŅŊÅúĘĩÄŋÉžŊâŧ°ûĒŧ¤ĩÄËÎīTUDCAĻÍģ×ĐĄĘķßMĐĐ×ĸÉäŖŦ×ĸÉäÖίēķĩÄÍģ×ĐĄĘķÔÚÄX˛ŋĐÎBēÍšĻÄܡŊÃæžųĶĐī@ÖøĩĸÄÉÆŖ¨Fig. S6ŖŠĄŖ
ČģļøŖŦąMšÜTUDCAĩÄÖίĘĮĶĐЧĩÄŖŦĩĢ×÷Õßī@Čģ˛ģÕJéß@ĘĮŌģˇNēÜĀíĪëĩÄÖίˇŊĘŊŖŦŌōéß@ĶĩÄÖίˇŊĘŊĐčŌĒģŧÕßÃŋĖėļŧ×ĸÉäTUDCAĄŖËųŌÔ×÷ÕßÔOĶÁËͨß^AAV-PHPeBŪD§Ō°ÉúĐÍWDR4ĩÄíŦFĻÍģ×ĐĄĘķßMĐĐÖίĩġŊˇ¨ŖŦAAV-PHPeBĻĐĄĘķÄX˛ŋĩĸĐȞЧÂĘēܸßŖŦÍŦrŌ˛ĩÄ´_ĘšĩÃÍģ×ĐĄĘķÔÚŊĶĘÜÖίēķŖŦÄX˛ŋĩÄĐÎBēÍĪāĒšĻÄÜĩÃĩŊÁËī@ÖøĩÄģÖÍŖ¨Fig. 6ŖŠĄŖ
×îēķŖŦ×÷ÕßĀûĶÃiPSCÕT§Ģ@ĩÃĩÄČËîÉņŊÔĒ¸Éŧ°ûŖ¨NSCsŖŠßMĐĐÁËķwÍâōŖŦˇÖeßMĐĐÁËTUDCAĖĀíēÍŌ°ÉúĐÍWDR4ŪD§ÖίŖŦŌÔÔušĀß@ɡNÖίˇŊĘŊĻČËîģŧÕßÅR´˛ÉĪŗŖŌÎģücÍģ×WDR4ĩÄЧšûēÍÔÚÅR´˛ĒĶÃrÖĩĄŖß@ɡNÖίˇŊĘŊļŧŋÉŌÔī@ÖøģÖÍWDR4Íģ×ĩÄČËîNSCsĩÄÔöÖŗËŽÆŊŖŦ˛ĸÔÚˇÖģ¯ĩÄß^ŗĖÖĐžŊâWDR4ÍģקÖÂĩÄĩōÍöŖ¨Fig. 7ŖŠĄŖ
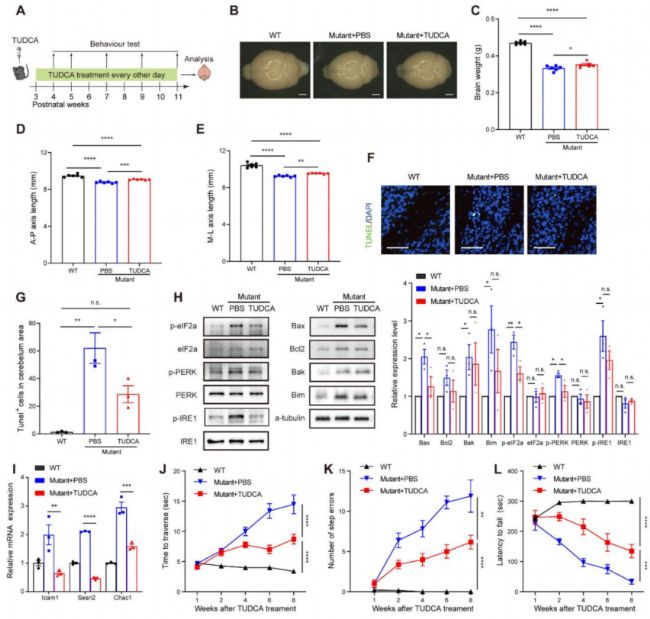
Fig. S6. TUDCAÖίtRNA m7GĐŪīĪÂÕ{ĐĄĘķĩÄÄXšĻÄÜ
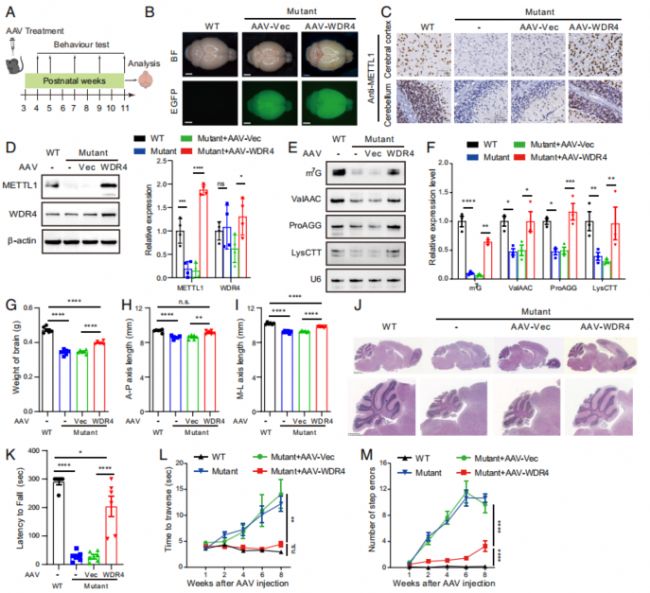
Fig. 6. tRNAĐŪīĪÂÕ{ĐĄĘķĩÄéLÆÚÖί¸ÉîA
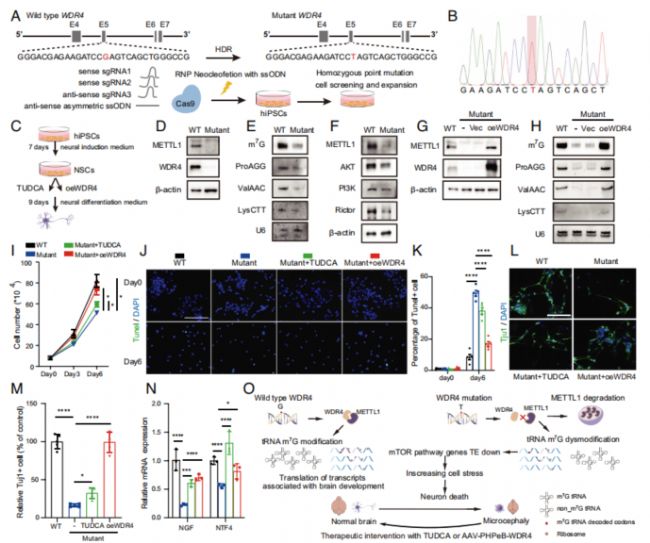
Fig. 7. TUDCAÖίģōģØŅaŌ°ÉúĐÍWDR4ĖáÉũ§ĶĐWDR4Íģ×ĩÄČËîiPSCíÔ´ĩÄNSC´æģîÂĘēÍšĻÄÜ
ŅĐžŋŋŊY
ąžÎÄĩÄÍęŗÉļČēܸßŖŦ×÷ÕßĩÄßŨžˇĮŗŖĮåÎúŖŦÄąíĐÍĩŊCÖÆÔŲĩŊģØŅaļŧĶĐÔŧÉîČëĀÖĩÄĖŊžŋŖŦËųĶĐĩÄōÔOĶēÍCÖÆĖŊË÷ļŧžožoúĀ@ÖøÅR´˛ÉĪĩÄąíĐÍÕšé_ŖŦß@Ō˛×ÎÄÕÂĩÄŊYÕžßĶиü´ķĩÄŋÉÄÜąģëHĒĶÃÔÚÅR´˛ÖίÖĐŖŦ˛ģëyŋ´ŗö×÷Õßąŗž°ÖĒ×RēÍōÔOĶˇŊÃæĩÄÄÜÁĻĪāŽÔúĄŖ
ąžÎÄÖĐĩÄsnRNA-seqÔÚąžžÍÍęÕûßBØĩÄšĘĘÂäęģųĩAÉĪå\ÉĪĖíģ¨ŖŦŗäˇÖ°l]ÁËÎŧ°ûŪDäŊMWĩÄ×îÖ÷ŌĒŨŖŦŧ´žĢ´_
^ˇÖŧ°ûÖŽégÔÚŪDäËŽÆŊÉĪĩÄŧÎĸ˛îŽŖŦ˛ĸͨß^ß@ˇN˛îŽĘ´_ĩØÕŌĩŊÁËŌģˇNžßĶĐī@Öø˛îŽĮŌÅcąíĐ͸ßļČÎĮēĪĩÄŧ°ûĐÍŖŦ×÷éąžÎÄšĘĘÂÖ÷žĩÄŅaŗäŖŦŌ˛ĘĮĪōÖøÁíŌģŅĐžŋŌŊĮÖ§žĩÄŗõ˛ŊĖŊË÷ĄŖŽČģŖŦąžÎÄĩÄsnRNA-seqĩūÖĐČÔČģĶĐÔSļāĐÅĪĸØŊ´ũÍÚžōēÍō×CŖŦģōÔSδíŋÉŌÔßMŌģ˛ŊĩØžĢĘĩØĖŊÃ÷WDR4Ļ×÷Õß°lŦFĩÄĖØļ¨ŧ°ûîĐÍ/ĐÍĩÄÕ{ŋØCÖÆŖŦģōÕßWDR4ĻÆäËûŧ°ûîĐÍ/ĐÍ´æÔÚĩÄÔÚÕ{ŋØĩČĩČŖŦß@ĐŠŅĐžŋĘĮŌģÖđu°lÕšĩÄß^ŗĖŖŦËųŌÔß@ĐŠžÍŌĒÆÚ´ũ×÷ÕßģōēķČËδíĩÄŅĐžŋÁËĄŖ

- ÉĖIģ¯ģųŌōēĪŗÉĩÄ°lÕšvŗĖĄĸÚ ŨēÍČËš¤ÖĮÄÜĐÂ
- RNA-BSŊŌĘžNONOĩ°°×ͨß^Õ{ŋØmRNAĩÄm5CĐŪīēÍŋÉ×ŧôĮĐ´ŲßMátWßMÕš
- WGBSŊŌĘžAtSAMSͨß^DNAŧ×ģųģ¯Ö˛Îīģ¨Æ÷šŲ°lĶũĩÄąíĶ^ßz÷CÖÆ
- ēĪŗÉÉúÎīWŖēÄ Ąą×xļŽÉúÃüĄą ĩŊ Ą°ÔėÉúÃüĄą ĩÄĩÚČũ´ÎÉúÎī¸īÃü
- m5C RNAŧ×ģųģ¯yĐōŖ¨m5C MeRIP-seqŖŠŧŧĐgŊéŊBÅcĒĶÃ
- RRBSĩȡÖÎöŊŌĘžÍŦÂŅëpÉú×ĶÅcÄIĐĄĮōVß^ÂĘĪāęPĩÄDNAŧ×ģųģ¯Îģüc
- WGBS+ChIP-seqŧŧĐgŊŌĘžCdx2ŪDäŌō×ĶÔÚ°lĶũÅcˇBÖĐĩÄĶBŊYēĪCÖÆ
- ēęģųŌōŊMWŊŌĘžØiŧSÅcÍÁČĀÖĐŋšžúËÎīŋšĐÔģųŌōÅcŊđŲŋšĐÔģųŌōĩÄš˛´æ
- 2025˛ŽēĀÉúÎī´ēŧžČĢøŅ˛Övé_ĸŖŦÔįøBķÃûÕŧĪČC
- ČA´ķÖĮÔė¸ąŋ˛ÃÖĐø ^ŋŊĀíÅíggŌģĐĐĩŊÔL˛ŽēĀÉúÎī
- ˛ŽēĀļĘÂéLŗöĪ¯ŖžĢĖØĐÂÖĐĐĄÆķI°lÕšŅ´ķÕ¯˛ĸ°lŅÔ
- ˛ŽēĀÉúÎīëp11ŋņgĖØģŨģîĶíŌuŖŦĪŪrĪŪÁŋ
- âųÃĀͨĩÂÍÆŗöŗÉąžrķwōÎŧ°ûyĐōŖ¨FFPEģōČĢŅĒŖŠ
- ˛ŽēĀÉúÎī2024ĪĩÁĐÅāĶ°āŖ¨ęÎ÷ÕžŖŠķÃûé_ĸ
- ˛ŽēĀÎŧ°ûFlexÄęÖĐžŪģŨŖŦ1.1W/ĶąžģîĶrŧ´ĸŊØÖš
- Ë{ž°ŋÆĐÅDAP-seqŧŧĐgĪāęPÎÄÕÂ6ßB°lŖŦŋIF 95.2


